
Chromosomen und Chromatin: Eine Frage der Verpackung
Chromosomen sind mehr als passive Träger der Erbinformation: Ihre Struktur hat großen Einfluss auf die Aktivität der Gene. So tragen sie dazu bei, die Identität und Funktion der Körperzellen festzulegen.
Menschliche Chromosomen – könnte man sie entrollen und hintereinander legen – bilden einen Faden von fast zwei Meter Länge. Der Zellkern, in dem sie ruhen, ist jedoch mikroskopisch klein: Sein Durchmesser beträgt etwa ein Hundertstel Millimeter. Das Erbgut muss daher auf kleinstem Raum verpackt werden1: Dazu werden die DNA-Fäden fein säuberlich auf winzige Spindeln gedreht.
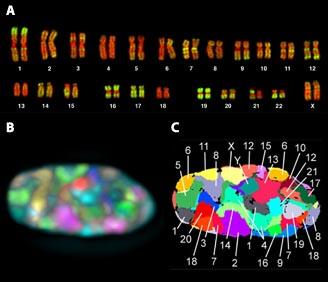
Im Zellkern herrscht also höchste Ordnung. Allerdings ist die DNA an manchen Stellen dichter gepackt, an anderen etwas lockerer. Dies schränkt den Zugang zum Erbgut ein. Die Packungsdichte der DNA dient also auch dazu, die Aktivität der Gene und die Funktion der Zelle zu steuern.
Inhalte
- Chromosom, Chromatin...
- Nukleosomen...
- Perlenkette...
- Engste Packung...
- Eu- und Heterochromatin...
- Signale...
- Mechanismen...
- Bedeutung...
Chromosom und Chromatin – wo liegt der Unterschied?
1879 fand der deutsche Anatom Walther Flemming einen Farbstoff, der auffällige Strukturen im Zellkern sichtbar machte. Diese Färbungen enthüllten jene X-förmigen Gebilde, die uns heute noch aus Schulbüchern vertraut sind. Aus dem griechischen Wort für Farbe – chroma – entstand schließlich der Begriff Chromosom.
Chromosomen nehmen die auffällige X-Form allerdings nur in einer bestimmten Lebensphase der Zelle ein – während der Zellteilung oder Mitose. Dann hat die Zelle ihr Erbgut gerade verdoppelt und muss es auf zwei Tochterzellen aufteilen. Um diese Aufteilung zu ermöglichen, wird die DNA „kondensiert‟: Ihr langer Strang wird auf möglichst kleinem Raum zusammengepackt. Die X-förmigen, kondensierten Chromosomen lassen sich dann problemlos trennen und fehlerfrei in zwei neue Kerne aufteilen.
Die Chromosomen sind nur während der Zellteilung unter dem Mikroskop zu sehen. Nach der Teilung gehen die Zellen wieder in ihren normalen Arbeitsmodus über – in der Fachsprache Interphase genannt. Die Chromosomen entfalten sich teilweise und füllen den Zellkern fast vollständig aus. Unter dem Mikroskop sind sie dann nicht mehr als eigene, isolierte Einheiten zu erkennen. Dieser aufgelockerte Zustand des Erbguts wird als Chromatin bezeichnet.
Die Begriffe Chromosom und Chromatin bezeichnen also zwei unterschiedliche Zustände des Erbguts. Während der Zellteilung sind die einzelnen Chromosomen in ihrer X-Form deutlich sichtbar. Im Arbeitsmodus der Zelle hingegen füllt das Erbgut als aufgelockertes Chromatin den Zellkern gleichmäßig aus.
Wie sind die Chromosomen und das Chromatin aufgebaut?
Das Speichermedium der Erbinformation ist die DNA: Ihre langen Ketten bestehen aus vier verschiedenen Buchstaben, deren Abfolge unter anderem die Baupläne der Proteine verschlüsselt. Sie ist damit der zentrale Bestandteil der Chromosomen und des Chromatins.
In der Zelle kommen die DNA-Moleküle jedoch nicht „nackt‟ vor: Winzige Protein-Spindeln wickeln kleine Stücke des DNA-Fadens auf. Diese Spindeln lassen sich zu größeren Gruppen vereinen, die dem Erbgut eine geordnete Struktur verleihen2.
Nukleosomen – kleine Spindeln für die DNA
Die Grundeinheit des Chromatins ist das Nukleosom: Es besteht aus jeweils acht Proteinen, die sich zusammenlagern und eine winzige Spindel erzeugen. Die Proteine tragen eine positive elektrische Ladung und ziehen daher die negativ geladene DNA elektrostatisch an. Die Spindel ist groß genug, um 147 Bausteine oder Basenpaare der DNA-Helix aufzuwickeln.
Nach einem kurzen Stück freier DNA – der sogenannten Linker-DNA, die beim Menschen ungefähr 50 Basenpaare lang ist – kann sich das nächste Nukleosom anlagern. Das Chromatin ähnelt dann einer langen Reihe von Perlen, die auf einer Kette aufgereiht sind.
Die Spindelproteine werden Histone genannt. Für den Aufbau einer Spindel werden vier verschiedene Histon-Varianten benötigt3: jeweils zwei Einheiten der Histone H2A, H2B, H3 und H4. Die Kernbereiche der Histone bilden den Körper der Spindel. Die beiden Enden der Histone ragen jedoch meist aus der Spindel heraus und können mit anderen Proteinen wechselwirken. Dabei können kleine Moleküle an die Histone angehängt werden, die wichtige Signale für die Struktur des Chromatins vermitteln.
Gefaltete Perlenketten
Die Grundstruktur des Chromatins – die Perlen auf der Kette – kann sich noch weiter verdichten. Eine wichtige Rolle spielt dabei eine weitere Histon-Variante: Das Histon H1 bindet an die Linker-DNA und sorgt dafür, dass die Nukleosomen dicht gepackt aneinander liegen. Der Durchmesser des Chromatin-Fadens verdreifacht sich dabei – von etwa 10 auf 30 Nanometer (ein Millionstel Millimeter).
Schließlich können weitere Gerüstproteine dafür sorgen, dass sich der dickere Faden noch weiter verknäuelt. Dabei entstehen zusammenhängende Gruppen, die Hunderte von Nukleosomen enthalten können4. Diese Bereiche haben häufig eine Größe von etwa 200 Nanometern2.
Verdichtet bis zur Sichtbarkeit
Wenn eine Zelle in die Teilungsphase eintritt, verdichtet sich das Chromatin weiter zu einer sehr kompakten Masse. Eine wichtige Rolle spielt dabei der Condensin-Komplex, ein Verbund aus mehreren Proteinen und Enzymen. Die einzelnen Chromosomen trennen sich nun voneinander und werden unter dem Mikroskop sichtbar.
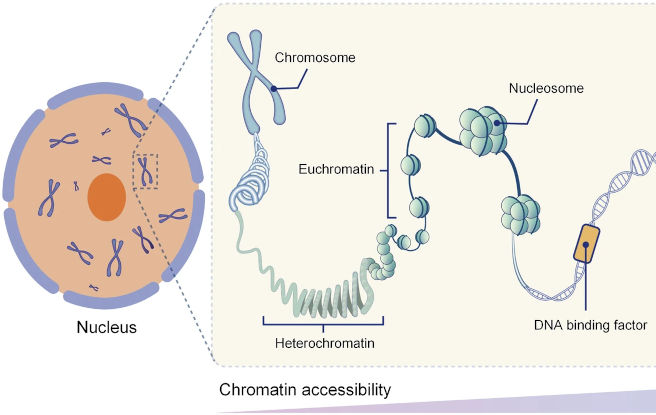
Was ist Euchromatin? Und was Heterochromatin?
Die Packungsdichte des Chromatins ist nicht gleichmäßig: Einige Bereiche sind sehr dicht gepackt, andere etwas lockerer. Diese beiden Formen nennt man Heterochromatin und Euchromatin. Ihre Verteilung ist nicht zufällig, sondern streng geregelt. Denn die Packungsdichte spielt eine wichtige Rolle für die Aktivität der Gene und die Funktion der Zelle.
Die dichtere Packung des Heterochromatins hängt damit zusammen, dass es in der Regel eine höhere Anzahl von Histon-Proteinen und Nukleosomen enthält. Vermutlich ist es deshalb auch für andere Proteine und Faktoren nur schwer zugänglich: Die Aktivität der Gene ist somit im Heterochromatin weitgehend abgeschaltet5.
Das Euchromatin enthält in der Regel weniger Nukleosomen und ist daher weniger dicht gepackt. In diesen Bereichen ist die DNA für wichtige Regulationsproteine – die Transkriptionsfaktoren – besser zugänglich. Diese Faktoren spielen eine zentrale Rolle bei der Entscheidung, welche Gene zu welchem Zeitpunkt angeschaltet werden. Das Euchromatin umfasst vermutlich nur etwa 3 Prozent des gesamten Erbguts5.
Die Unterschiede in der Packungsdichte sind vermutlich nicht sehr groß: Heterochromatin ist nach Schätzungen nur etwa 1,5-mal dichter als Euchromatin. Neben der Anzahl der Nukleosomen gibt es vermutlich noch weitere Faktoren, die die Zugänglichkeit der genetischen Information bestimmen2,4.
Wie wird der Aufbau von Chromatin gesteuert?
Das Erbgut enthält keine festen Regeln, welche seiner Teile als Euchromatin und welche als Heterochromatin vorliegen sollen. Diese Aufteilung kann sich langsam herausbilden, wenn sich der Körper und seine Gewebe entwickeln. Oder sehr schnell, wenn eine Zelle auf besondere Herausforderungen antworten muss.
Der Auf- und Umbau von Chromatin ist ein komplizierter Prozess, an dessen Steuerung viele Signale und Schaltkreise mitwirken. Und die praktische Umsetzung erfolgt oft durch große Protein-Komplexe, die aus mehreren Enzymen bestehen5.
Welche Signale lösen den Umbau des Chromatins aus?
Eigenheiten des Erbguts, Veränderungen im Stoffwechsel, Reize aus der Umwelt – es gibt viele Signale, die die Struktur des Chromatins beeinflussen. Proteine und RNA-Moleküle können diese Informationen in den Zellkern weiterleiten und den Umbau in Gang setzen
Andere Signale hinterlassen Spuren direkt am Chromatin. Damit beschäftigt sich das Gebiet der Epigenetik – mit dem Erbgut verbundene Informationen, die nicht in der Abfolge der DNA-Buchstaben verschlüsselt sind. Wichtige epigenetische Signale sind die Histon-Modifikationen und die DNA-Methylierung5.
- Histon-Modifikationen: Die freien Enden der Histon-Proteine können verändert werden, wenn besondere Enzyme kleine Moleküle daran anhängen. Diese sogenannten Modifikationen können die Eigenschaften der Histone verändern oder Signale an andere Proteine übermitteln. Sie bilden ein kompliziertes Muster, das für die Zugänglichkeit der DNA eine wichtige Rolle spielt.
- DNA-Methylierung: Die DNA kann ebenfalls chemisch verändert werden: Enzyme koppeln ein kleines Molekül an den Strang, das als Methyl-Rest bezeichnet wird. Diese Markierung findet sich häufig in Bereichen des Chromatins, die nicht leicht zugänglich sind.
- Nicht-codierende RNA: Dabei handelt es sich meist um kleinere RNA-Moleküle, die nicht zur Herstellung von Proteinen dienen. Sie können dazu beitragen, den Zugang zur der DNA zu steuern.
- Transkriptionsfaktoren: Diese Proteine steuern meist die Aktivität von Genen. Einige von ihnen sind aber auch in der Lage, sich an dicht verpacktes Chromatin zu binden und es für andere Transkriptionsfaktoren zugänglich zu machen4.
Welche Proteine sind am Umbau beteiligt?
Der Umbau von Chromatin bedeutet häufig, dass Nukleosomen aufgebaut oder verschoben werden. Teile der Chromatin-Fäden können auch zu Schleifen geformt werden, um ihre Zugänglichkeit zu erhöhen. An diesen Prozessen sind zahlreiche Proteine und Enzyme beteiligt. Eine wichtige Rolle spielen unter anderem:
- Chromatin-Remodeling-Komplexe: Mehrere Enzyme bilden einen zusammenhängenden Verbund, um Nukleosomen einzubauen oder zu verschieben. Dabei verbrauchen sie Energie in Form von ATP. Vier große Familien dieser Komplexe sind im Zellkern aktiv5.
- Cohesin: Dieser große Verbund aus mehreren Enzymen hat die Form eines Rings: Der DNA-Strang kann dort eingefädelt und in einer Schleife angeordnet werden. Cohesin spielt auch eine wichtige Rolle beim Zusammenhalt der Chromosomen während der Zellteilung6.
Warum ist die Struktur des Chromatins so wichtig?
Gene können nur aktiv werden, wenn besondere Proteine – die Transkriptionsfaktoren – an die DNA binden und das Ablesen der Buchstaben (die Transkription) einleiten. Viele Transkriptionsfaktoren können sich aber im Zellkern nicht frei bewegen: Sie gelangen nicht in Bereiche, die zu dicht oder aus anderen Gründen unzugänglich sind. Die Struktur des Chromatins bestimmt also mit, an welche DNA-Sequenzen die Transkriptionsfaktoren andocken können5. Und damit auch, welche Gene in einer Zelle angeschaltet werden.
Epigenetische Markierungen, die die Struktur des Chromatins stark beeinflussen, bleiben oft auch nach einer Zellteilung erhalten. Dies kann dazu beitragen, die Identität einer Zelle festzulegen: Eine Leberzelle wird immer Leberzellen hervorbringen, eine Muskelzelle immer Muskelzellen. Es gibt aber auch Hinweise darauf, dass die Struktur des Chromatins die Entwicklung von Lebewesen und die Bildung von Körpergeweben beeinflusst7. Allerdings sind hier noch viele Fragen offen – die Forschung steht oft noch am Anfang.
2 Li et al., Phase-separated chromatin compartments: Orchestrating gene expression through condensation, Cell Insight, Dezember 2024 (Link)
alle Referenzen anzeigen
3 Nho und Kim, Dynamics of nucleosomes and chromatin fibers revealed by single-molecule measurements, BMB Reports, Januar 2025 (Link)4 Fillot und Mazza, Rethinking chromatin accessibility: from compaction to dynamic interactions, Current Opinion in Genetics & Development, Februar 2025 (Link)
5 Chen et al., Chromatin accessibility: biological functions, molecular mechanisms and therapeutic application, Signal Transduction and Targeted Therapy, Dezember 2024 (Link)
6 F. Uhlmann, A unified model for cohesin function in sisterchromatid cohesion and chromatin loop formation, Molecular Cell, März 2025 (Link)
7 Cardona et al., Bridging spatial and temporal scales of developmental gene regulation, Current Opinion in Genetics & Development, März 2025 (Link)
Aufbau des Erbguts
Epigenetik
Genomforschung
⇒ Aufbau des Erbguts
- Chromosomen und Chromatin mehr...
- Nicht-codierende DNA mehr...
- Das Genom als RNA-Maschine mehr...
- Das ENCODE-Projekt mehr...
- Der genetische Code mehr...
- Evolution des genetischen Codes mehr...
- Single Nucleotide Polymorphism (SNP) mehr...
⇒ Wissenswertes
⇒ Epigenetik
Kurz und knapp
- lange Abschnitte der DNA-Fäden des Erbguts sind auf kleinen Spindeln aufgerollt
- diese Nukleosomen können noch enger verpackt und verdichtet werden
- während der Zellteilung ist der DNA/Protein-Verbund stark gefaltet und unter dem Mikroskop als Chromosom sichtbar
- in aktiven Zellen ist der DNA/Protein-Verbund etwas lockerer und füllt als Chromatin den Zellkern aus
- die Struktur des Chromatins hat einen direkten Einfluss auf die Aktivität der Gene